Спектр совместимых растворимых веществи их распространение у
микроорганизмовСтраница 1

Спектр совместимых растворимых веществ, обнаруженных у прокариот и эукариот, весьма широк и разнообразен (Roberts, 2004; 2005). Термин “совместимые растворимые вещества” был предложен для соединений, которые не ингибируют метаболические процессы, но защищают клетку и клеточные компоненты в условиях водного стресса (Brown, 1976). Этот термин применим к органическим осмолитам, которые предохраняют макромолекулы от ингибирующего действия неорганических ионов или органических молекул. Некоторые органические вещества могут также защищать клетки и макромолекулы при замораживании-оттаивании, высушивании и воздействии высокой температуры (da Costa et al., 1998). Так, например, в опытах in vitro показано, что эктоин стабилизирует лактатдегидрогеназу и другие ферменты при воздействии высушивания, высокой и низкой температуры. Благодаря этим свойствам, осмолиты могут использоваться в косметике и биотехнологии (Ventosa and Nieto, 1995).
Осмопротекторы относятся к различным классам органических соединений, специфических для разных групп галофильных и галотолерантных микроорганизмов. Проведенный скрининг осмопротекторов у более чем 200 галофильных изолятов, включая цианобактерии (Reed et al., 1984), аноксигенные фототрофные бактерии (Trüper and Galinski, 1986), аэробные хемогетеротрофы, протеобактерии a- и g-подклассов (виды рода Halomonas, Vibrio, Pseudomonas), актиномицеты (виды Actinopolyspora, Nocardiopsis), бациллы и родственные виды Staphylococcus и Salinicoccus, умеренно галотолерантные виды Brevibacterium и Corynebacterium (Bernard et al., 1993; Frings et al., 1993), позволил разделить эти соединения на следующие основные группы (Galinski, 1995):
- полиолы (глицерин, арабит, манит), сахара и их производные (сахароза, трегалоза и глюкозилглицерин);
- цвиттерионные соединения и бетаины (глицинбетаин);
- аминокислоты (пролин, глутамин и глутамат);
- N-ацетилированые диаминокислоты (ацетилорнитин, ацетиллизин);
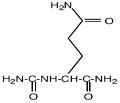
- амидные производные глутамата (N-карбамоилглутамиламид);
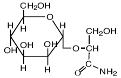
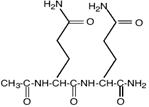
- эктоины (эктоин, гидроксиэктоин);
- метилированые сульфосоединения (диметилсульфониопропионат), накапливающиеся у цианобактерий и морских водорослей.
Эти исследования привели к ряду важных обобщений:
1. Синтезируемые de novo полиолы, такие как глицерин, арабит, инозит, часто накапливаются у галофильных и галотолерантных грибов, устойчивых к солям растений, но не обнаружены у галофильных бактерий.
2. Все “совместимые вещества”, синтезируемые или транспортируемые из среды, накапливаются в концентрациях, превышающих 500 мМ, и являются полярными, хорошо растворимыми молекулами, не несущими суммарного заряда.
3. Заряженные аминокислоты, такие как глутамат, b-глутамат, бетаинглутамат и другие, не накапливаются в очень больших количествах (более 400 мМ).
Осмолиты могут быть разделены на основании затрачиваемой клеткой энергии для их биосинтеза (Oren, 1999). При этом наиболее энергетически выгодным является биосинтез глицерина, эктоина и глицин-бетаина, наименее выгоден синтез сахарозы и трегалозы. Многие бактерии способны накапливать одновременно ряд осмопротекторов, при этом преобладание того или иного осмолита во многом определяется энергетическим статусом клетки и доступностью источника азота.
Все вышеперечисленные группы осмолитов можно разделить на три химические категории: 1) анионные растворимые вещества, 2) незаряженные растворимые вещества и 3) цвиттерионые растворимые вещества. В таблице 1 приведены распространенные у микроорганизмов совместимые растворимые вещества.
|
Распространение у микроорганизмов |
литература | ||
|
Анионные органические осмолиты, содержащие карбоксильную группу | |||
|
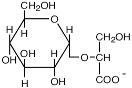
α-Глутамат |
|
E. coli; Halomonas elongata; Methanohalophilus portucalensis; Methanobacterium thermoautotrophicum; Natronococcus occultus; Halobacterium sp.NRC-1 и H. salinarum; Erwinia chrysanthemi а также многие галотолерантные бактерии и метаногены |
Roberts, 2004 Goude et al., 2004 |
|
β-Глутамат |
|
Methanothermococcus thermolithotrophicus; Methanocaldococcus jannaschii; Methanotorris igneus; Nocardiopsis halophila, у всех видов Nocardiopsis, Thermotoga neapolitana и T. maritime |
Galinski and Trüper,1994; da Costa et al., 1998; Martin et al., 1999 |
|
Глутаматбетаин |
|
Calothrix sp. N181 |
Mackay et аl., 1984 Roberts, 2004 |
|
α-Гликозил-глицерат |
|
Agmenellum quadruplicatum; Stenotrophomonas maltophilia; Methanohalophilus portucalensis Erwinia chrysanthemi |
Roberts, 2004 Goude et al., 2004 |
|
α-Маннозил-глицерат |
|
Dehalococcoides ethenogenes; Methanothermus fervidus; Pyrococcus furiosus; Rhodothermus marinus и R. obamensis; Methanohalophilus portucalensis; Thermus thermophilus; Dehalococcoides ethenogenes; Rubrobacter xylanophilus; Aeropyrum pernix; Pyrococcus sp.; Thermococcus sp.; Archaeoglobus fulgidus |
Empadinhas et al., 2004 Santos and da Costa, 2002 Martins et all., 1999; Goncalves et all., 2003 Lamosa et all., 1998; Neves et all., 2005 |
|
Анионные органические осмолиты, содержащие фосфатные или сульфатные группы | |||
|
Ди-мио-инозит-1,1’-дифосфат |
|
Archaeoglobus fulgidus; Methanotorris igneus; Pyrococcus furiosus и P. woesei; Pyrodictium occultum; Thermotoga maritima |
Martin et al., 1999; Roberts, 2004 Santos and da Costa, 2002 Martins et all., 1997 Martins et all., 1996 |
|
α-Диглицерин- фосфат |
|
Archaeoglobus fulgidus |
Martins et all., 1997 Goncalves et all., 2003 |
|
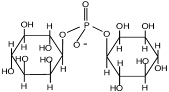
Цикло-2,3-дифосфоглицерат |
|
Methanobacterium thermoautotrophicum; Methanopyrus kandleri; Methanothermus fervidus |
Martins et all., 1997 |
|
Сульфотрегалоза |
|
Natronococcus occultus; Natronobacterium sp. |
Desmarais et al., 1997 |
|
Незаряженные органические осмолиты | |||
|
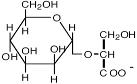
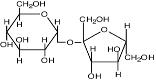
Гликозил-глицерин |
|
Морские и пресноводные цианобактерии: Synechocystis sp., Microcystis firma; фототрофные эубактерии: Rhodobacter sulfidophilum; Pseudomonas mendocina и P. рseudoalcaligenes; Stenotrophomonas sp. |
Reed et al., 1986 Galinski, 1995; da Costa et al., 1998 |
|
α-маннозил-глицерамид |
|
Rhodothermus marinus и R. obamensis |
Silva et al., 1999 |
|
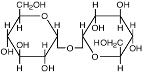
Трегалоза |
|
Pyrobaculum aerophilum; Sulfolobus solfataricus и S.ambivalens; Thermoproteus tenax; Thermoplasma acidophilum; Actinopolyspora halophila; Scytonema species cемейства Thiocapsa, Thiocystis, Chromatium, а также Azotobacter chroococcum, Klebsiella pneumoniae, Pseudomonas aeruginosa, Rhizobium meliloti, Chromohalobacter israelensis; Desulfovibrio halophilus; Rhodothermus obamensis; Narialba magadii T. Thermophilus; Sinorhizobium meliloti |
Galinski, 1995; da Costa et al., 1998 Page-Sharp et al., 1999 Nunes et all., 1995; Elbein et all., 2003 Goufii et all., 1999 |
|
Сахароза |
|
Synechocystis sp., Anabaena sp., Phormidium autumnale; Scytonema species; Chroococcidiopsis sp. Sinorhizobium meliloti |
Reed et al., 1986; Hershkovitz et al., 1991; Page-Sharp et al., 1999; Desplats et all., 2005. |
|
Na-карбамоил-L-глутамин-1-амид |
|
Ectothiorhodospira marismortui |
Oren et al., 1991. |
|
N-α-ацетил-глутаминил-глутаминамид |
|
Sinorhizobium meliloti; Rhizobium leguminosarum; Pseudomonas aeruginosa; пурпурные сульфобактерии Rhizobium meliloti, P. fluorescens, P. aeruginosa, P. putida, P. mendocina |
Smith and Smith, 1989; D’Souza-Ault et al., 1993; Kets et al., 1996. |
|
Манносахароза |
|
Agrobacterium tumefaciens |
Smith et al., 1990 |
|
Цвиттерионые органические осмолиты | |||
|
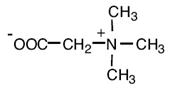
Глицинбетаин |
|
Thioalkalivibrio versutus; Actinopolyspora sp.; Actinopolyspora halophila; Halorhodospira halochloris; Methanohalophilus portucalensis FDF1; Methanosarcina thermophila; Methanothermobacter marburgensis; Synechococcus sp. DUN52, B. subtilis E. coli, Staphylococcus xylosus Sinorhizobium meliloti, Halomonas elongata Methanobacterium thermoautotrophicum Marburg Ectothiorhodospira halochloris, Actinopolyspora halophila, галотолерантный фототроф Aphanothece halophytica Methanohalophilus portucalensis; Listeria monocytogenes |
Lai et al., 1991 Boch et al., 1996, Østerås et al., 1998, Canovas et al., 2000, Nyyssölä et al., 2000, Waditee et al., 2003 Bayles et all., 2000 |
|
Эктоин |
|
Sporosarcina pasteurii; Bacillus pasteurii; Marinococcus halophilus; Brevibacterium epidermis, B. linens; Thioalkalimicrobium aerophilum; Vibrio cholerae и V.costociola; Chromohalobacter israelensis и C. salexigens; Halorhodospira halochloris; Halomonas elongata H. israelensis, H. variabilis; метилотрофы: Methylomicrobium alcaliphilum, Mm. keniense, Mm. buryatense; Methylophaga alcalica и M. natronica, Methylarcula terricola и M. marina; и многие галотолерантные бактерии |
Canovas et al., 1997; Wohlfarth et al., 1990 Khmelenina et al., 1999 Doronina et all., 2000; Doronina et all., 2003a, b |
|
Гидроксиэктоин |
|
Halomonas elongata; Chromohalobacter salexigens; Nocardiopsis halophila; N. dassonvillei; Micrococcus halobius; Marinococcus halophilus, Marinococcus sp. M52; Brevibacterium casei, B. iodinum, B. linens, Streptomyces chrysomallus, S. griseolus; Salibacillus salexigens |
Roberts, 2004 Frings et al., 1993 Severin et al., 1992 |
|
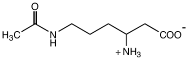
Nε-ацетил-β-лизин |
|
Methanothermococcus thermolithotrophicus; Methanosarcina thermophila, M. mazei Gö1, M. acetivorans. M. barkeri; Meyhanococcus jannaschii и M. maripaludis; Methanohalophilus portucalensis FDF1; Halobacillus halophilus Methanosarcina thermophila, Methanogenium cariaci, Methanohalophilus sp. и Methanococcus sp. Sporosarcina halophila, Planococcus citreus |
Pflüger et al., 2003 Martin et al., 1999; Roberts, 2004 da Costa et al., 1998 |
|
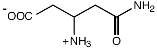
β-глутамин |
|
Methanohalophilus portucalensis FDF1 |
Roberts, 2004 |
|
Пролин |
B. subtilis и Planococcus citreus, Azospirillum brasilense некоторых видов Staphylococcus и Salinicoccus |
Kempf et all., 1998 Miller et all., 1996 | |
|
Диметил-сульфонио-пропионат |
|
Морские водоросли; поглощается из среды E. Coli, S. typhimurium |
Galinski, 1995; Welsh, 2000 Cosquer et. al., 1999; Pichereau et al., 1998 |
|
Пипиколиновая кислота |
|
Brevibacterium ammoniagenes, Sinorhizobium meliloti |
Gouesbet et al., 1992 Gouffi et al., 2000 |
|
Карнитин |
|
Поглощаются из среды: E. coli, Lactobacillus plantarum, Listeria monocytogenes, Pediococcus halophilus, Brevibacterium linens, Pseudomonas aeruginosa, Listeria monocytogenes |
Sleator, Hill, 2001 Kets et al., 1994 Bayles et all., 2000 Jebbar et al., 1998 |
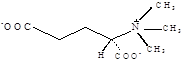
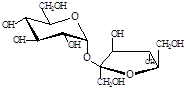
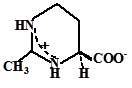
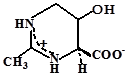
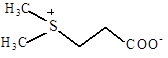
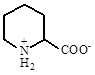
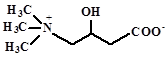
Также смотрите:
Лиганды
Великое множество лигандов, описанных в научной литературе, можно разбить на две главных категории. Это лиганды с индивидуальной специфичностью и лиганды с групповой специфичностью, В качестве индивидуальных лигандов могут выступать ферменты, а также субстраты или инг ...
Ламаркизм
Ж.Б. Ламарк, ботаник при Королевском ботаническом саде, первый предложил развернутую концепцию эволюции органического мира. Он остро осознавал необходимость формулирования новых теоретических целей, методологических установок биологического познания; потребность в обо ...
Учет и выпуск молоди
Целесообразно использовать сплошной объемный метод учета молоди. Его применяют на рыбоводных заводах при выпуске молоди из прудов, площадь которых не превышает 2—4 га. Учет количества выращенной молоди осуществляют в рыбоуловителе, изготовленном из металлической сетки ...