Физиологические эффекты гормонов, плохо проникающих в клеткуСтраница 1

Гормоны, плохо проникающие в клетку (белково-пептидные, катехоламины), соединяются с рецепторами на клеточной мембране и вызывают внутриклеточные эффекты через вторые посредники.
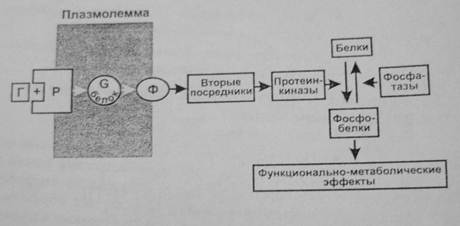
Большинство гормонов после присоединения к рецепторам активируют регулируемый гуанилнуклеотидами G-белок мембраны. Активация состоит в обмене ГДФ на ГТФ в α-субъединице G-белка. При этом происходит его распад на α-субъединицу и βγ-димер, имеющие самостоятельные регуляторные эффекты. Более изучено действие α-субъединицы, являющейся ферментом ГТФазой. Гидролиз ГТФ до ГДФ освобождает энергию, используемую для передачи регуляторного сигнала. ГДФ-α-субъединица соединяется с βγ-димером, вновь образуя ГДФ—G-белок. Имеется несколько видов G-белков (GS, Gi ,Gq ,GO), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (GS), так и ингибировать (Gi) образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.
В действии гормонов, плохо проникающих в клетки, можно выделить 4 системы: аденилатциклазную, фосфоинозитидную, гуанилатциклазную и тирозинкиназную.
Аденилатциклазная система. Эту систему используют гормоны, осуществляющие свой эффект через аденилатциклазу и синтез второго посредника—циклического АМФ (3,5-цАМФ). Действие гормона может, как активировать, так и тормозить его образование.
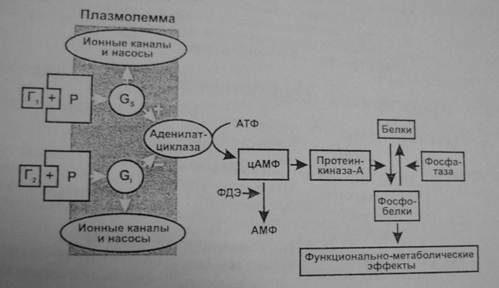
- Комплекс «гормон + рецептор» активирует GS –белок, который активирует аденилатциклазу и образование цАМФ (например, так действуют адреналин через β1/β2-рецепторы, АДГ через V2-рецепторы, глюкагон, АКТГ, ТТГ, ФСГ, ЛГ, МСГ, кортиколиберин, соматолиберин, кальцитонин, хориогонадотропин человека, паратгормон).
- Комплекс «гормон + рецептор» активирует Gi-белок, который тормозит аденилатциклазу и образование цАМФ (например, так действуют адреналин через α2-рецепторы, ангиотензин II через АТ-рецепторы, аденозин через А1-рецепторы, соматостатин, опиоиды через μ-, δ- и κ-рецепторы, ацетилхолин через М2-холинорецепторы гладкомышечных клеток).
- цАМФ влияет на функции клетки через изменение активности протеинфосфокиназы-А, фосфорилирующей различные клеточные белки: мембранные (изменяется ее проницаемость), ферменты (изменяется метаболизм клетки), ядерные (изменение транскрипции), рибосом (изменение трансляции), микротрубочек (двигательные эффекты, изменение формы клетки) и др.
Фосфоинозитидная система. Эту систему используют гормоны, которые, действуя через Gq-белок, активируют в мембране фосфолипазу С.
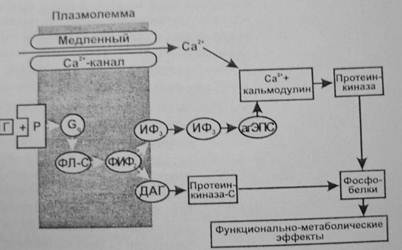
Она образует из фосфолипида мембраны (фосфатидилинозитолдифосфата) два посредника—инозитол—1,4,5—трифосфат (ИФ3) и 1,2—диацилглицерол (ДАГ).
- ИФ3, переходя в цитоплазму, действует на рецепторы гладкой ЭПС, что приводит к открыванию Са2+-каналов и выходу Са2+ в цитозоль. Са2+ при этом выступает в качестве третьего посредника, который связывается с белком кальмодулином и активирует Са2+-кальмодулинзависимую протеинкиназу, фосфорилирующую эффекторные белки, влияющие на функции клеток, их пролиферацию и дифференцировку (регулирует активность аденилатциклазы и синтез цАМФ, фосфолипазы—А2 и синтез простагландинов, тромбоксана, простациклина, гуанилатциклазы и синтез цГМФ, фосфодиэстеразы и разрушение цАМФ и цГМФ; участвует в регуляции секреции инсулина, гормонов щитовидной железы, гипофиза и надпочечников, кишечной секреции и секреции нейромедиаторов). (Роль Са2+ как посредника действия гормонов обусловлена его сильным влиянием при низких концентрациях 10-8—10-6 М на метаболизм и функции клетки. Гормоны могут увеличивать концентрацию Са2+ в цитозоле как в результате открывания Са2+-каналов клеточной мембраны, так выхода Са2+ из гладкой ЭПС. В клетке есть специальные белки, связывающие Са2+ при низкой концентрации—кальмодулин, гуанилатциклаза, тропонин С и др. Присоединение Са2+ к этим белкам изменяет их конфигурацию и свойства. Это приводит к изменению многочисленных функций клеток—метаболической, секреторной, сократительной, биоэлектрической и др.)
- ДАГ, оставаясь на внутренней поверхности мембраны, активирует протеинкиназу С, фосфорилирующую эффекторные белки (например, активирует киназу легкой цепи миозина), а также синтез простагландинов из арахидоновой кислоты, что изменяет функции клетки.
- Через ИФ3 и ДАГ действуют адреналин (α-рецепторы), вазопрессин (V1), ацетилхолин (М1), ангиотензин II (AT1), гистамин (Н1), серотонин (5-НТ2), холецистокинин, окситоцин, ВИП и др.
Также смотрите:
Многоножка-камнелаз
Многоножка-камнелаз, или костянка (Lithobius forficatus L.), принадлежит к отряду однопарноногих (Chilopoda) в классе многоножек (Myriapoda).
Когда мы отворачиваем куски коры старых пней с целью отыскания различных животных, нередко перед нашими глазами промелькнет с ...
Гносеологическая стрела
Свое название эта стрела получила от «гносеологии» - науки о познании. Человек начал познавать мир почти одновременно со своим появлением и познает его довольно интенсивно. Сложность методик и полученных результатов растут очень быстро. ...
Модификационная изменчивость в теории эволюциию Дарвинизм
В 1859 году Чарльз Дарвин опубликовал свою работу на эволюционную тему под названием «Происхождение видов путём естественного отбора, или сохранение благоприятных рас в борьбе за жизнь». В ней Дарвин показал постепенное развитие организмов как результат естественного ...